This article was published in Scientific American’s former blog network and reflects the views of the author, not necessarily those of Scientific American
Chemists studying metabolism in living organisms usually classify it into two kinds; primary and secondary. Primary metabolism is concerned with the production and reactions of essential biomolecules like proteins, sugars and lipids. Secondary metabolism refers to the production of small molecules which, although not essential, are still important in a variety of key functions.
Secondary metabolism is especially pronounced in plants. Since plants unlike animals cannot move around, they have evolved specialized strategies to protect themselves and breed. One of the most successful strategies is to employ secondary metabolism to produce a bewildering array of small molecules that are used in almost every imaginable function, from defense against predators to establishing symbiotic relationships with bacteria to attracting pollinating agents. Two of the most important kinds of secondary metabolites are alkaloids and terpenes, both of which have been used by humans for centuries as drugs, fragrances, pesticides and food. Alkaloids include strychnine, morphine and caffeine and terpenes include menthol, cholesterol and vitamin A. Antibiotics are also secondary metabolites. Needless to say, the impact of these molecules on human commerce and culture has been enormous.
A gallery of secondary metabolites found in key culinary ingredients, including capsaicin (peppers), allicin (garlic) and menthol (mint)
On supporting science journalism
If you're enjoying this article, consider supporting our award-winning journalism by subscribing. By purchasing a subscription you are helping to ensure the future of impactful stories about the discoveries and ideas shaping our world today.
Another reason why secondary metabolism is important is because it has shed light on key physiological processes in human beings. For instance, the biosynthetic pathways and enzymes that produce cholesterol in humans are very similar to those producing the steroid ergosterol in fungi. The enzymes involved in these pathways often perform a spectacular, elegant dance in which multiple chemical bonds are formed with exquisite timing and selectivity. Comparison of the amino acid sequences of these enzymes across different species can provide valuable information about their evolutionary relationships. Such relationships reaffirm that evolution seeks common solutions to diverse problems.
One of the questions that arises when studying the evolutionary history of secondary metabolism is how its enzymes evolved separately from those orchestrating primary metabolism. In a recent perspective in Science (1), a group from the Salk Institute hypothesizes that gene duplication might have played a preponderant role in the evolution of these proteins. The rationale here comes from the kinds of reactions that enzymes catalyze. Enzymes are usually very specific for a given substrate; in fact organic chemists would give anything to achieve that kind of specificity in their laboratory syntheses. But it turns out that the enzymes for secondary metabolism are not as specific as their primary counterparts; they can bind to many different kinds of molecules and transform them into different products. In terminology which surprisingly enough has become a standard part of the scientific literature, these enzymes are promiscuous.
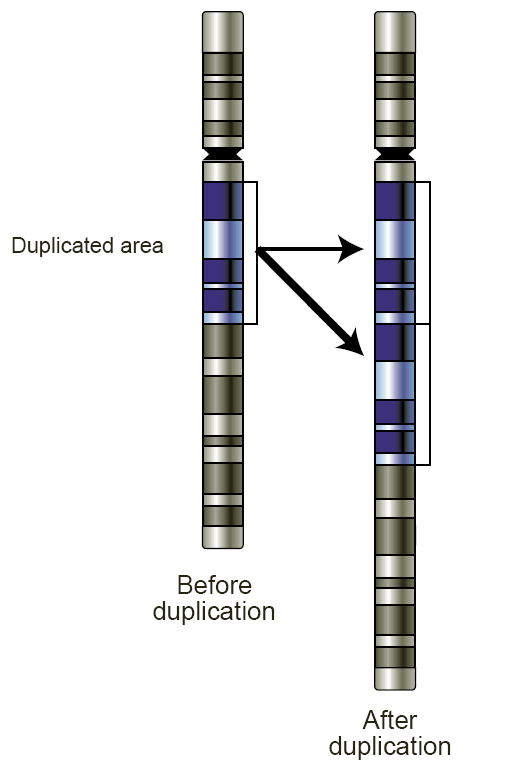
How could this broad specificity have come about? Usually when evolution 'fixes' a function in a gene for a protein, it is difficult to tamper with that function without introducing deleterious and sometimes catastrophic effects. The genetic tool that makes such risky experimentation possible constitutes one of evolution's great inventions - gene duplication. Gene duplication has proven to be a potent strategy to increase the information content in the genome and enable diversity of function. The principle is simplicity itself. At some point a gene undergoes duplication as a random event. This event frees up one of the copies to undergo mutations without jeopardizing the original function since the other copy is untouched and continues going about its business. Through successive evolution and natural selection, the second copy often ends up mutating into something that performs a novel and hitherto unexplored function. Now there's every possibility that the changes might be deleterious and would make the gene extinct. But the ones that matter are those that stick. Because of the sheer number of mutations that evolution subsumes, there's always going to be some that improve the function; once this happens natural selection takes over and retains those changes.
{kind=link}
The blood clotting cascade and the proteins involved in it; many of these arose from a common ancestor partly through gene duplication (Image: John Lynch)
{kind=link}
Duplicated genes can undergo mutations - including deleterious ones - without affecting the original function (Image: Wikipedia Commons)
One of the best examples of gene duplication is in the evolution of blood clotting enzymes. There's more than a dozen of these, and a comparison of their sequences and genes across species indicates that they must have started from a common ancestor with modest blood clotting abilities. Successive duplication events allowed this gene to proliferate into a whole family of clotting enzymes, each with its own substrate specificity. Another example from the same fold is the evolution of the globin family of genes, with hemoglobin carrying oxygen in the bloodstream and myglobin binding it in the muscles. The construction of a whole family of proteins for a critical system like blood clotting or transport demonstrates the power of gene duplication.
In the case of secondary metabolism, the authors postulate that the enzymes must have undergone duplication and then diversified. A typical scenario has a gene A for an enzyme catalyzing the reaction of substrate X undergoing duplication. The new gene is now free to tinker and undergoes mutations. Many of these are deleterious or neutral, but at some point a single mutation or a combination of them creates a new gene B whose protein product can now bind to both substrates X and Y; the protein has become promiscuous. This will be true especially if Y has a valuable role in the metabolism of the organism and therefore exerts positive pressure for a particular mutation resulting in gene B. If you study the enzymes responsible for secondary metabolism, you find that many of them are in fact promiscuous, either in the same creature or in different ones. A particularly appealing example is that of a protein called dihydroflavonol reductase (DFR), which depending on the organism it's working in produces either red or blue pigments from two different substrates. And what goes for multiple substrates also goes for the multiple end products of enzymatic reactions; in a striking case, an enzyme called TEAS acts on a single substrate to generate one major and no less than 23 minor products. The exact functions of promiscuous enzymes will depend on the species, environmental factors and further genetic events but gene duplication seems to be the most plausible general mechanism here.
{kind=link}
Compounds called anthocyanins which are responsible for the colors of flowers - pansies in this case - are made by enzymes which can be the products of gene duplication (Image: Commons)
However as humans well know from painful experience, promiscuity comes at a price. In this case the price that these enzymes pay for their molecular dalliances is a loss of activity. As the authors describe, these enzymes typically bind their molecular partners 30 times less tightly than the enzymes of primary metabolism. This principle is general; enzymes often trade efficiency for specificity. And although the article does not say it, I wonder if this behavior also affects the flexibility of the enzymes and makes them floppier in order to accommodate more diverse substrates. It's almost as if being unfaithful to one substrate affects their performance. There may be a lesson in there somewhere.
Notwithstanding the suggestive analogies, this is an intriguing hypothesis and seems entirely consistent with what we know about evolution. Evolution has developed many simple mechanisms for exploring diversity and increasing the number of molecular players in its grand biochemical orchestra. Gene duplication has been one of the most successful strategies. Next time you sip a cup of coffee, watch a wound heal or admire the blue or red color of flowers, think of secondary metabolism, and thank those promiscuous enzymes and their duplicate genes.
References:
1. The Rise of Chemodiversity in Plants, Science, 2012: 1667-1670
2. Jiang and Doolittle, Proc. Nat. Acad. Sci. USA, 2003, 100, 7527